Publication Information
Frequency: Continuous
Format: PDF and HTML
Versions: Online (Open Access)
Year first Published: 2019
Language: English
| Journal Menu |
| Editorial Board |
| Reviewer Board |
| Articles |
| Open Access |
| Special Issue Proposals |
| Guidelines for Authors |
| Guidelines for Editors |
| Guidelines for Reviewers |
| Membership |
| Fee and Guidelines |
 |
Optimizing the Sex Ratio and Oviposition Material for Mass Production of Sycanus Collaris (f.) (Hemiptera: Reduviidae)
Tewee Maneerat1* & Pornsawan Poopat1
1Agricultural Innovation and Management Division, Faculty of Natural Resources, Prince of Songkla University, Hatyai, Songkla, 90110, Thailand
Received Date: March 11, 2021; Accepted Date: March 18, 2021; Published Date: March 26, 2021;
*Corresponding author: Tewee Maneerat, Agricultural Innovation and Management Division, Faculty of Natural Resources, Prince of Songkla University, Hatyai, Songkla, 90110, Thailand. Email: tewee.m@psu.ac.th
Citation: Maneerat T, Poopat P (2021) Optimizing the sex ratio and oviposition material for mass production of sycanus collaris (f.) (hemiptera: reduviidae). Adv Agri Horti and Ento: AAHE-143.
Abstract
Biological control of insect pests by releasing natural enemies relies on stable mass production of natural enemies. In Thailand, Sycanus collaris (F.) (Hemiptera: Reduviidae) has been mass-multiplied and released to control leaf-eating caterpillars and small beetles in the forests and crops. To mass produce S. collaris for release, there were gaps in knowledge about the process. This study aimed to determine the most suitable sex ratio of S. collaris which produced the maximum number of offsprings using yellow mealworm, Tenebrio molitor (L.) (Coleoptera: Tenebrionidae) as diet and to test oviposition materials that simplify egg collection. Colonies with adult sex ratios of 1:1, 1:2, 1:3, 1:4, and 1:5 (male: female) were reared separately to compare egg quantity, quality, and adult lifespan. The results showed no significant differences in number of eggmass, number of eggs per eggmass, percentage hatchability, longevity of male and female, and oviposition period (P > 0.05). Sex ratio of 1:3 was chosen due to lower cost of production, enough males in the container, and consistency of eggs produced. To reduce the cost of rearing while considering egg production over time, S. collaris adults should be reared up to 54 days. Colony with sex ratio 1:3 was tested with oviposition materials (craft paper, black, green, and clear plastic) in rearing containers, and found that craft paper was the most preferred material (P < 0.01). Number of eggs per eggmass and percentage hatchability were not different among materials (P > 0.05).
Keywords: Assassin Bug; Biocontrol; Mass Production; Oviposition Material; Sex Ratio
Introduction
Sycanus collaris (F.) (Hemiptera: Reduviidae) is one of interesting insect predators to use in a biological control program. Its distribution is mostly found in South Asia and Southeast Asia, including Thailand (Ambrose, 2006; Jamjanya et al., 2014; Sahayaraj, 2012).
S. collaris was documented to consume a number of forest and agricultural pests, such as, Helicoverpa armigera, Mestocharella indica, Hyblaea puera, Blepharipa zebina, Spodoptera litura, and Spodoptera exigua in Asia including Thailand (Jamjanya et al., 2014; Sahayaraj, 2012). S. collaris in nymphal and adult stages feed by piercing on their prey with proboscis and sucking hemolymph out of the prey. S. collaris was shown to be an effective predator under laboratory and field conditions and was suggested that this predator should be mass- reared and used as a biocontrol agent (Jamjanya et al., 2014; Rajan et al., 2017).
To produce enough insect predators for release and colony maintenance, a well- established rearing system is necessary. Successful mass production systems need an efficient management in limiting resources, namely, space, labor, and time, so that the cost of production remains affordable. Certain rearing conditions, such as, sex ratio and egg collection substrate, which produce the maximum number of predators should be investigated. None of the studies in mass rearing of predatory Hemipteran insects investigated the effect of sex ratio on number of offspring produced and adult lifespan. In Rhynocoris fuscipes and Prithesancus plagipennis (Hemiptera: Reduviidae) mass rearing, adults were typically reared in 1:5 (male:female) (Grundy et al., 2000; Tomson et al., 2017). The advantage of such ratio was that less adults (or less male adults) were maintained in the rearing container and thus reduced the cost of rearing. In S. collaris, not much was known about the recommended sex ratio in mass rearing system.
Egg collection in mass rearing can be a challenge in terms of high maintenance, skill requirement, and insect acceptance to specific oviposition sites. Heteropteran females were reported to exhibit a sequential of oviposition site selection events (Lundgren, 2011). Suitable habitat was needed to be located before egg oviposition began. Then, a microhabitat (i.e. plant or other substrates) was selected. Chemical or morphological cues might be required to locate the microhabitat. Lundgren (2011) further discussed that proximate cues were more important for females to select suitable sites over unsuitable ones. Many Heteropteran species accepted abiotic oviposition sites (i.e. cage and paper towel) under laboratory conditions, although plants were primary oviposition sites in the field (Lundgren, 2011). For example, cage and paper towel were chosen as oviposition sites more than plants in Geocoris punctipes (Hemiptera: Geocoridae) (Lundgren, 2011; Silva and Panizzi, 2009). Little was known about S. collaris oviposition materials which could be used in the rearing.
Considering available resources, two aspects of S. collaris mass production process were investigated, with aims to propose a more affordable and efficient rearing process. The objectives of this study were to 1) test the effect of sex ratio on number of offspring and adult lifespan and 2) test oviposition materials that simplified egg collection and maintained egg quality.
Materials and Methods
The experiment was conducted under laboratory conditions (27 ± 3 C, 55-80 % RH, and 12L: 12D photoperiod) at the Faculty of Natural Resources, Prince of Songkla University, Hatyai Campus, Songkla, Thailand.
Stock culture of sycanus collaris (f.) And tenebrio molitor (l.)
S. collaris colony was obtained from Biological Control Laboratory, Prince of Songkla University, Songkla, Thailand, and were reared in a plastic container (24.5 cm- diameter, 11 cm-height) with screen. Non sibling S. collaris males and females were allowed to mate in the containers to avoid inbreeding. Eggs were collected to rear in a separate container. First to second instar nymphs were fed with T. molitor pupae and filtered water using wet cotton balls. Thrid to fifth instar and adults fed on mixed sizes of T. molitor larvae. An orange jessamine branch with leaves, Murraya paniculata, was provided in the rearing container of first to third instars and a plastic basket (17 cm-diameter, 5 cm-height) was placed in the rearing container of fourth to fifth instars and adults to increase surface area and provide microhabitats for the insects.
S. molitor eggs to adults were reared in reared plastic containers (24.5 cm-diameter, 11 cm-height) with screen and fed with a mixture of wheat bran, corn bran, rice bran, yeast, and calcium. Each stage was maintained separately. Pupae were left in the rearing container until adult emergence. Adults were fed with the same diet as larvae and they were allowed to mate and lay eggs on wax paper. Eggs were collected every 3 days. Frass was sifted through a fine screen weekly to allow more access to diet.
Effect Of Sex Ratio On Egg Quantity And Quality
To examine the effect of parental sex ratio on egg quantity and quality, colonies of newly emerged adults with sex ratios of 1:1, 1:2, 1:3, 1:4, and 1:5 (male:female) were maintained separately in clear plastic containers (13 cm-diameter, 5 cm-height) with orange jessamine branch with leaves. Since one male was included per container, this study eliminated the effect of male mating competition. The experiment was a completely randomized design (CRD) and had five replications (one replication represented one container). Two yellow mealworm larvae per one adult were given daily from adult emergence to death. Adult longevity (time from emergence to death), number of eggmass, number of eggs per eggmass, hatchability (total number of nymphs hatched per total number of eggs laid), and oviposition periods (time between the first eggmass to the last eggmass by a female) of the predators were recorded every 24 hours. This study was conducted in April 2019 and ended when adults were dead.
Effect of Oviposition Materials on Egg Quantity And Quality
To determine which oviposition material was the most suitable, 5 freshly molted males and 15 females (1 male: 3 females) were placed in a rearing container (24.5 cm- diameter, 11 cm-height). The experiment used CRD and had five replications (one replication represented one container with four types of oviposition materials). Oviposition materials (one of each), paper roll, black plastic roll, green plastic roll, and transparent plastic roll (11 cm-length, 4 cm-width, 4 cm-height) (Figure 1) were added to the rearing container after 14 days because oviposition began 14-16 days after adult emergence. Females located and laid eggs on any of four materials including rearing container. The adults were given 30 T. molitor larvae (2-3 g) three times weekly.
Figure 1: Oviposition materials, paper roll, black plastic roll, green plastic roll, and transparent plastic roll (11 cm-length, 4 cm-width, 4 cm-height) inside a rearing container. 
Oviposition rolls were removed to collect egg and new rolls were replaced weekly until 60 days-post experiment, since the sex ratio study revealed that eggs from over 54 days- old females were low in number and hatchability. In addition, egg-laying frequency per female was every 7-10 days and egg incubation period was 16-17 days. Number of eggmasses on oviposition rolls and container, eggs per eggmass, percentage hatchability, number of dead males, and females were recorded daily. This study was conducted during September-November 2019.
Statistical Analysis
To compare the effect of sex ratio, number of eggmass per female, number of eggs per eggmass, hatchability, male longevity and female longevity were analyzed by one-way analysis of variance using SPSS (version 23.0; SPSS Inc.; Chicago, IL, USA). Similarly, number of eggmass on oviposition materials, number of eggs per eggmass, and hatchability were compared by one-way ANOVA and Duncan’s multiple range test (DMRT).
Results and Discussion
Effect of sex ratio on egg quantity and quality
All colonies with different sex ratios produced eggmasses in the range of 3.47±0.90 – 4.42±1.28 eggmasses per female, 52.19±16.66 – 77.11±3.64 eggs per eggmass, and hatchability of 51.36±25.97 – 58.93±14.38% (Table 1). There were no significant differences in number of eggmass laid by a female (F4, 18 = 0.70, P = 0.60), number of eggs per eggmass (F4, 18 = 2.36, P = 0.092) and hatchability (F4, 18 = 0.093, P = 0.98) across all treatments. (mean ± sd) 2/ns = no significant differences (P > 0.05)
Sex ratios (male: female)
N
No. Eggmass/female (mean ± sd)1/
No. Eggs/eggmass (mean ± sd)
Hatchability (%)
Male
Female
1:1
5
5
4.25 ± 1.26
52.19 ± 16.66
51.36 ± 25.97
1:2
5
10
4.42 ± 1.28
73.64 ± 13.25
55.82 ± 17.56
1:3
5
15
3.62 ± 0.97
65.62 ± 12.10
54.28 ± 17.14
1:4
5
20
4.17 ± 0.52
77.11 ± 3.64
58.93 ± 14.38
1:5
5
25
3.47 ± 0.90
70.14 ± 10.32
52.53 ± 8.03
F test
ns2/
ns
ns
1/Mean ± SD within the same column analyzed by one-way analysis of variance
No significant differences in male longevity (F4, 18 = 1.77, P = 0.18), female longevity (F4, 18 = 1.06, P = 0.40) and oviposition period (F4, 18 = 1.58, P = 0.22) were observed among five sex ratios (Table 2). 2/ns = no significant differences (P > 0.05)
Sex ratios (male: female)
N
Male longevity (d) (mean ± SD)1/
Female longevity (d) (mean ± SD)
Oviposition period (d) (mean ± SD)
Male
Female
1:1
5
5
40.80 ± 12.32
46.60 ± 21.05
29.50 ± 7.00
1:2
5
10
46.50 ± 11.78
62.92 ± 19.15
44.00 ± 11.01
1:3
5
15
34.00 ± 7.42
53.81 ± 21.12
43.86 ± 15.56
1:4
5
20
49.00 ± 7.55
65.50 ± 25.01
53.33 ± 18.01
1:5
5
25
48.67 ± 14.57
62.87 ± 18.98
43.33 ± 8.62
F test
ns2/
ns
ns
1/Mean ± SD within the same column analyzed by one-way analysis of variance
Number of eggs produced by a female changed as it aged. Colony with 1:1 showed high number of eggs per female on days 14-35 and 42 and egg number dropped sharply after that (Figure 2A). Colony with 1:2 showed high number of eggs per female on days 30, 38, and 42, and egg number began to slowly decline (Figure 2B). Colony with 1:3 showed high number of eggs per female on day 19-25 and it began to decline slowly (Figure 2C). Colony with 1:4 showed high number of eggs per female on days 17, 34, and 41 and it began to decline slowly (Figure 2D). Colony with 1:5 showed high number of eggs per female on day 16 and 29 and it began to drop sharply (Figure 2E). Adult sex ratios of 1:2, 1:3, or 1:4 may be suitable to maximize S. collaris offspring’s. Sex ratios of 1:1 and 1:5 showed low number of eggs per female (Figure 2A; Figure 2E) and hatchability over time. After 54 days, egg numbers were low in all sex ratios.
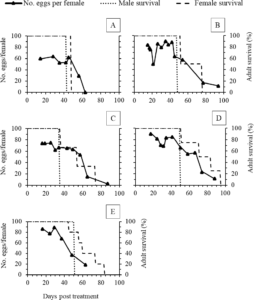
Figure 2: Changes of egg number per female Sycanus collaris (F.), male survival (%) and female survival (%) when reared in colonies with sex ratios of 1:1 (A), 1:2 (B), 1:3 (C), 1:4 (D) and 1:5 (E) were shown over time (days post treatment) under laboratory conditions (27± 3°C, 55-80 % RH).
No report on the effect of sex ratio on offspring in Hemipteran was found in the literature. Thus, studies on this topic that could be related to the results were of other insect orders. The results of this study were similar to a study by Damiens et al. (2019). The authors reported no significance in egg production when reared Aedes albopictus (Diptera: Culicidae) in 1:1, 1:3, and 1:5 (male:female). The authors observed an increase in egg production as the number of females increased. Alternatively, another study in Anastrepha ludens (Diptera: Tephritidae) found the significant effect of sex ratio on egg production, hatchability, and female survival probabilities when compared 1:4 to 1:1 (male:female) (González-López et al., 2019). Both studies (Damiens et al., 2019; González-López et al., 2019) at least showed the similar trend that as the number of females increased, the egg production also increased. However, in the current study, colonies of 1:2, 1:3, and 1:4 showed slightly higher egg production and hatchability than of 1:1 and 1:5.
Sex ratio may influence number of offspring and adult lifespan. Although the results from this study did not support that sex ratio had an impact on adult lifespan, the study on A. ludens demonstrated that ratios with more females tended to reduce males and female lifespan (González-López et al., 2019). However, Desa et al. (2018) suggested that more males in the rearing cages lowered adult lifespan because male mating competition required energy and possibly caused stress and mortality. This study eliminated the effect of male competition on adult lifespan. During the experiment, there were temperature and humidity fluctuations that could affect egg hatchability and adult longevity, since these factors could cause unpredictability in insect populations over time (Chown and Terblanche, 2009; Sørensen et al., 2012).
It was also important to note that S. collaris female lifespan sometimes lasted longer than its oviposition period regardless of sex ratio, and studies also supported that fecundity in Hemipterans decreased as females were older (Javahery, 1994; Lundgren, 2011). To reduce the cost of rearing while considering egg production over time (Figure 2), S. collaris adults should be reared up to 54 days.
As the number of females increases in the rearing container, cost of production, such as amount of prey provided and handling time, will be reduced. On the contrary, the tradeoff of lowering number of males is an increase in risks of not having enough males to mate, since some males may die over time, thus results in unfertilized eggs. Besides, more males may ensure genetic variability in the colony. When sex ratios of 1:2, 1:3, and 1:4 were compared, the production cost of 1:4 would be the lowest, but this ratio had highest possibility of not enough males per container. Ratio of 1:3 would be more cost efficient than of 1:2 due to higher egg production per container. Overall, sex ratio of 1:3 (male:female) was recommended for S. collaris because it still lowered the production cost and the risk of insufficient males was not as high as more female-biased ratios. Number of eggs produced was also more consistent. Colonies of 1:3 should also be maintained for up to 54 days since egg production was low after this period. Additionally, it was worth repeating the experiment in a different time of year to compare if there was an effect of seasonality on number of offspring produced.
Effect of oviposition material on egg quantity and quality
S. collaris females laid eggs in clusters and attached eggmass on the surface of the materials provided. During oviposition, females started laying eggs from outer to inner part of the eggmass, and newly laid egg was attached vertically to previously laid eggs and the material surface. Secretion used by female to attach eggs and oviposition site was white and after a few days it dried out and appeared transparent yellow. Eggmass shapes could be hexagon, indistinct, or one row of eggs. Eggmasses laid on craft paper were mostly in hexagon shape (Figure 3A), whereas those found on all colors of plastic were variable in shapes (i.e. indistinct and row of eggs) (Figure 3B-D). Eggmasses on rearing container could be in hexagon (Figure 3E) or indistinct shape.
Among all materials placed inside a rearing container, females laid highest number of eggmass (46.00±10.58) on craft paper (F4, 20 = 53.89, P < 0.01). A rearing container was second most preferred by females (17.40±7.73 eggmass). Green, clear, and black plastic were laid 1.60±2.07, 1.00±1.73, and 0.20±0.45 eggmass, respectively. Different oviposition materials did not affect number of eggs per eggmass on craft paper (69.06±4.71), green plastic (69.70±11.60) and container (66.54±5.43) (F4, 11 = 0.71, P = 0.60). Similarly, different materials did not affect percentage hatchability of eggs laid on craft paper (73.22±9.29), green plastic (73.76±25.09) and container (66.54±5.43) (F4, 11 = 0.94, P = 0.47) ().

Figure 3: Oviposition materials, craft paper, black plastic, green plastic and clear plastic were placed inside a rearing container of 5 Sycanus collaris (F.) males and 15 females (1:3) under laboratory conditions (27 ± 3 C, 55-80% RH). Eggmasses laid on craft paper were mostly in hexagon shape (A), whereas those found on all colors of plastic were variable in shapes, specifically, indistinct (B-C) and a row of eggs (D). Eggmasses on rearing container could be in hexagon (E) or indistinct shape. 2/ Females from less than 3 replications laid eggs on black plastic and clear plastic (N < 3) and the data were excluded from number of eggs per eggmass and percentage hatchability comparisons.
Oviposition materials
No. of eggmass
No. of eggs/eggmass
Hatchability (%)
N
mean±SD
N
mean±SD
N
mean±SD
Paper
5
46.00±10.58a1/
5
69.06±4.71
5
73.22±9.29
Black plastic
5
0.20±0.45c
12/
68.00
1
97.06
Green plastic
5
1.60±2.07c
3
69.70±11.60
3
73.76±25.09
Clear plastic
5
1.00±1.73c
2
79.50
2
67.43
Container
5
17.40±7.73b
5
66.54±5.43
5
71.07±8.86
F test
**
ns
ns
1/ Mean±SD within the same column were analyzed by one-way analysis of variance. Different letters indicate significant difference (P < 0.05) using Duncan’s multiple range test (DMRT). ** = highly significant (P < 0.01), ns = no significant difference (P> 0.05).
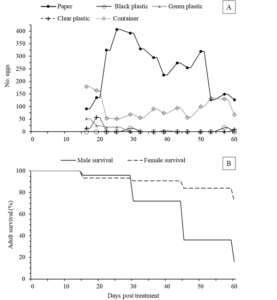
Figure 4: Oviposition materials, specifically, craft paper, green plastic, clear plastic, black plastic, and container were placed inside a rearing container of 5 S. collaris males and 15 females (1:3). Total number of eggs laid on each oviposition material (A) and adult survival percentage (B) were shown over time (N = 5).
Number of eggs laid on all materials was shown over 60 days (A). Egg number laid on craft paper reached its peak during days 23-29, then started to decline and increased again during days 46-50. Egg number laid on black, green, and clear plastic were low (mostly less than 50 eggs). Egg number laid on rearing container showed highest number on day 16, then continued to decrease and began to increase again on day 50-53 before dropping afterward. As for adult survival, males lived shorter than females (B). Male survival percentage dropped from 72% to 36% during days 31-46. Female survival percentage decreased more slowly. After 60 days, 72% of females were alive while only 16% of males were alive. Male lifespan in this study was longer than male lifespan (1:3 colony) in sex ratio study, but female lifespan was similar.
In S. collaris, females highly preferred craft paper over other materials. Without any oviposition materials, eggs were laid on container surface, such as lid and sidewall. A similar observation was found in Coleomegilla maculate (Coleoptera: Coccinellidae) (Riddick et al., 2019). Muslin gauze covered on container lid was used as an oviposition substrate in mass rearing of P. plagipennis (Grundy et al., 2000). Although there was no evidence about S.collaris oviposition site selection in the literature, it appeared from the present study that sites with air ventilation and ability to adhere to eggmass were hypothesized to explain why S. collaris chose craft paper and rearing container lid as oviposition sites rather than plastic ones under laboratory conditions.
One replication represented one container with all four materials. Craft paper, the most preferred oviposition materials, had good characteristics for use to collect S. collaris eggs. S. collaris eggmass was attached firmly to paper, which could be cut and transferred to another rearing container or glued to packaging. For plastic, females did not prefer to lay eggs on plastic rolls and rather laid eggs mostly on container lid (near screen). Eggmass on all colors of plastic did not remain adhered to the materials and fell off easily when secretion dried off. Eggmass laid on container lid could be difficult to remove, especially when the secretion adhered to the screen (part of the lid). Removing process by scraping sometimes damaged eggs. Moreover, the surface area of rearing container without any materials tended to be limited when 60 adults were maintained in one container. Adding materials would increase surface area and possibly reduce stress and competition for microhabitat. From this experiment, craft paper was the best oviposition material for S. collaris. Future study may test other variations of paper that differ in texture and color, because certain types of paper may be more cost-efficient to produce S. collaris.
Conflict of Interest: The authors declare that there are no conflicts of interest.
Acknowledgements: The authors would like to thank Ms Jusmin Niyomdecha for laboratory assistance. Facilities and financial support from Agricultural Innovation and Management Division, Faculty of Natural Resources, Prince of Songkla University are gratefully appreciated.
References
- Ambrose DP (2006). A checklist of Indian Assassin Bugs (Insecta: Hemiptera: Reduviidae) with taxonomic status, distribution and diagnostic morphological characteristics. Zoos’ Print J. 21:2388-2406.
- Chown SL, Terblanche JS (2009). Physiological diversity in Insects. Adv Insect Phys 33:50-152.
- Damiens D, Marquereau L, Lebon C, Le Goff G, Gaudillat B, et al. (2019). Aedes albopictus adult medium mass rearing for sit program Insects 10:1-9.
- Desa G, Tsegaye M, Lelisa K, Argiles R, Lema B, et al. (2018). Optimizing the sex ratio to maximize the yield of sterile males in Tsetse mass-rearing colonies. J. Entomol. 11:59-65.
- González-López GI, Solís-Echeverría E, Díaz-Fleischer F, Pérez-Staples D (2019). When less is more: Sex ratios for the mass-rearing of Anastrepha ludens (Diptera: Tephritidae). Econ. Entomol. 112:2997-3001.
- Grundy PR, Maelzer DA, Bruce A, Hassan E (2000). A mass-rearing method for the assassin bug Prithesancus plagipennis (Hemiptera: Reduviidae). Biol. Control 18:243-250.
- Jamjanya T, Siri N, Phanphinit S (2014). Life history of assassin bug, Sycanus collaris (Hemiptera: Reduviidae) and its efficacy to control insect pests. Khon Kaen Agric. J. 42: 130-135.
- Javahery M (1994). Development of eggs in some true bugs (Hemiptera-Heteroptera). Part I. Can. Entomol. 126:401-433.
- Lundgren JG (2011). Reproductive ecology of predaceous Heteroptera. Biol. Control 59:37-52.
- Rajan SJ, Suneetha N, Sathish R (2017). Biology and predatory behaviour of an assassin bug, Sycanus collaris (Fabricius) on rice meal moth, Corcyra cephalonica (Stainton) and leaf armyworm, Spodoptera litura (Fabricius). Agric. 12:1181-1186.
- Riddick EW, Wu Z, Eller FJ, Berhow MA (2019). Potential of 2, 4-dihydroxybenzoic acid as an oviposition stimulant for mass-reared ladybird J. Insect Sci. 19:1-6.
- Sahayaraj K, (2012). Artificial rearing on the nymphal developmental time and survival of three reduviid predators of Western Ghats, Tamil J. Biopestic. 5:218-221.
- Silva FAC, Panizzi AR (2009). Oviposition of the neotropical brown stink bug Euschistus heros (Heteroptera: Pentatomidae) on artificial and on natural Florida Entomol. 92:513-515.
- Sørensen JG, Addison MF, Terblanche JS (2012). Mass-rearing of insects for pest management: Challenges, synergies and advances from evolutionary physiology. Crop 38:87-94.
- Tomson M, Sahayaraj K, Kumar V, Avery PB, McKenzie CL, et al. (2017). Mass rearing and augmentative biological control evaluation of Rhynocoris fuscipes (Hemiptera: Reduviidae) against multiple pests of cotton. Pest Manag. Sci. 73:1743-1752.